Размер геномов на протяжении эволюции судьбы на Земле возрастал ускоренными темпами

Число генов у кукурузы вдвое превышает число генов у человека, а у червяка приблизительно такое же, как у человека.
Растолковать данный феномен ученые пока не смогут. Рисунок с сайта bio.fizteh.ru
Русские ученые на базе продуманной методики доказали, что рост размеров генома связан с повышением сложности организмов в ходе эволюции. Данный, казалось бы, очевидный вывод до сих пор не поддавался статистическому доказательству. Но, как продемонстрировали американские эксперты, данный рост не удается растолковать с помощью существующих популяционных накопления дрейфа и моделей генов случайных мутаций.
Диапазон изменчивости размеров генома огромен: он образовывает практически 7 порядков — от внутриклеточных бактерий с геномами в 150–200 тысяч пар нуклеотидов до некоторых бактерий с их 700 млрд пар нуклеотидов. Казалось бы, повышение сложности генома должно быть связано со сложностью морфологической организации. Бактерии несложны — им и несложного генома хватает, а дабы руководить развитием сложного животного, требуется более сложная генетическая «программа».
Но, как ни страно, подтвердить это предположение не получалось (см. C-value enigma). Берем ли мы безвыборочный массив видов либо выбираем отдельные филогенетические линии, не удается статистически подтвердить связь между размером геномов и уровнем организации.
К примеру, сходные в морфофизиологическом смысле виды злаков имеют 20-кратный разброс в размерах генома. В то же время у человека геном приблизительно в 40 раз меньше, чем у двоякодышащей рыбы Protopterus aethiopicus (см.: Comparison of different genome sizes).
Это несоответствие возможно дать добро двумя методами. Первый — доказать, что обстоятельства отсутствия, казалось бы, очевидной корреляции искусствен , то имеется связаны с методическими просчетами. Второй путь — доказать, что указанной связи вправду нет, и предложить другую содержательную обстоятельство трансформаций размера генома на протяжении эволюции.
В текущей научной литературе показалось два сообщения, помогающих разобраться в данной проблеме.
Одно из них предлагает новый методический подход к оценке связи размера генома и сложности организации (либо уровня эволюционного развития). Данный подход был создан группой русских экспертов под управлением А. В. Маркова из Палеонтологического университета (Москва) и размещён в «Палеонтологическом издании». Второе, размещённое в PLoS Genetics, разрешает осознать, как на большом растоянии продвинулись ученые, выбравшие второй путь.
Марков с сотрудниками продемонстрировали наличие связи между минимальным размером и уровнем организации генома представителей этого уровня — но не для всех живых организмов в целом, а лишь для одной эволюционной линии, ведущей от прокариот к млекопитающим. В данной эволюционной линии тенденция к росту сложности организма (отнюдь не являющаяся общим эволюционным законом) проявилась самый ярко. Специфика этого изучения в том, что в качестве меры размера генома в той либо другой группе организмов использован минимальный геном, а не диапазон, не средние величины и не случайные виды.
Наряду с этим внутриклеточные паразиты не рассматривались, поскольку они вовсю пользуются генами хозяина, экономя личные ресурсы. Конкретно минимальный геном в многочисленной группе организмов может оказать помощь примерно оценить количество нужной (неизбыточной) генетической информации, снабжающей существование представителей таксона. Более правильную оценку взять тяжело, потому, что нет полностью надежных способов отличить вправду ненужные участки ДНК от функционально значимых (к примеру, от некодирующих последовательностей, делающих регуляторные функции).
Не считая минимального генома для каждого эволюционного уровня учитывалось кроме этого примерное время появления данной группы организмов (см. таблицу).
| Уровень организации | Минимальный геном (млн пар нуклеотидов) | Примерное время появления (млн лет назад) | Вид — обладатель самого мелкого генома (не считая внутриклеточных паразитов) |
| 1. Всё живое | 1,3 | 4000 | Морская свободноживущая бактерия Pelagibacter ubique, штамм HTCC1062 |
| 2. Эукариоты | 9,2 | 2000 | Сумчатый гриб Ashbya gossypii — обладатель самого мелкого генома среди свободноживущих эукариот |
| 3. Животные (Metazoa) | 19,6 | 1250 | Нематода Pratylenchus coffeae |
| 4. Хордовые | 68,6 | 575 | Аппендикулярия Oikopleura dioica, представитель подтипа Tunicata (оболочники) типа хордовых |
| 5. Позвоночные | 342 | 540 | Костная рыба Tetraodon fluviatilis |
| 6. Тетраподы | 931 | 375 | Лягушка Limnodynastes ornatus |
| 7. Амниоты | 951 | 315 | Фазан Phasianus colchicus |
| 8. Млекопитающие | 1695 | 220 | Летучая мышь Miniopterus schreibersi |
| 9. Приматы | 2215 | 65 | Мартышка-прыгун Callicebus torquatus |
Ученые подобрали функции (размер генома в зависимости от времени появления), наилучшим образом аппроксимирующие эти сведенья. Оказалось, что эту зависимость оптимальнее обрисовывает гиперэкспоненциальная функция, то имеется степенная экспонента; несколько хуже — экспонента (см. рис. 1).
![]()
Рис. 1. Зависимость минимального размера генома от времени появления группы организмов.
По оси абсцисс отложено время появления, а по оси ординат — минимальный размер генома. Рисунок из обсуждаемой статьи в «Палеонтологическом издании»
Гиперэкспоненциальный рост говорит о том, что повышение генома — это самоускоряющийся процесс. Чем больше геном, тем выше скорость увеличения. Какие конкретно процессы лежат в базе этого автокатализа?
По предположению ученых, одним из источников повышения генома возможно формирование новых генов и регуляторных участков методом дупликации и перекомбинирования имеющихся функциональных участков. Много генов допускает и больший комплект вероятных перекомбинаций их участков, и эта функция, разумеется, не прямая, а степенная. Сложность сама собой порождает рост сложности.
Полученные эти заставляют обратить внимание на следующий факт. Самый резкий скачок в повышении минимального размера генома зафиксирован в позднем докембрии и раннем палеозое (600–540 млн лет назад), а большие скорость увеличения биоразнообразия, то имеется сложность более больших иерархических уровней судьбы, были достигнуты довольно много позднее — в финише мезозоя и в кайнозое (начиная приблизительно со 100 млн лет назад). Не исключено, что рост генома в какой-то мере содействовал росту биоразнообразия, предоставив дополнительные степени свободы для генетических комбинаций (не смотря на то, что нужно не забывать, что взрывной рост разнообразия в кайнозой и мезозой происходил в главном за счет относительно легко устроенных животных, таких как насекомые и моллюски, и в меньшей степени за счет костистых рыб, млекопитающих и птиц).
Это расхождение должно заинтересовать будущих исследователей.
Так, в случае, если осмысленно доходить к выбору последовательности усложняющихся форм и оценкам размера геномов, то удается не только подтвердить связь между сложностью организации и геномом, но и заметить черты самоускоряющегося процесса. Это, непременно, макроэволюционный масштаб преобразований. Наряду с этим в пределах каждого таксона рост генома может и не выявляться, а также его вовсе может и не быть.
Так, в эволюции птиц и прокариот преобладающей тенденцией, по-видимому, было уменьшение, а не повышение размера генома.
Во второй статье американские исследователи из университета Райса (Хьюстон) и Калифорнийского университета в Риверсайде постарались отследить микроэволюционные процессы, регулирующие рост генома. В данном случае анализировалась не «нужная» часть генома, которую возможно примерно оценить по минимальному размеру генома в пределах многочисленной группы, а целый геном полностью, включая его «факультативную» часть, размер которой может сильно варьировать, к примеру, из-за размножения «эгоистических» мобильных элементов и иных повторяющихся последовательностей. Предполагается, что размер генома должен быть связан обратной зависимостью с численностью популяции. В солидных популяциях за счет вероятностных процессов слабовредные мутации должны действенно отсеиваться отбором.
Считается, что большая часть дупликаций повторяющихся участков ДНК являются именно слабовредные мутации. Исходя из этого в громадных популяциях отбор обязан удерживать геном от разбухания, а в мелких, наоборот, слабовредные дупликации и новые копии мобильных элементов будут накапливаться, увеличивая размер генома.
Контролировать эту модель направляться с громадной осторожностью, поскольку исследователя поджидают неожиданные статистические трудности. В зависимости от характера ветвления филогенетического древа выборки получаются различные результаты. В случае, если смешать филогенетически далекие и родные ветви, то получается «средняя температура по поликлинике»: смешанная выборка демонстрирует якобы прямую зависимость, а любая отдельная ветвь может характеризоваться обратной зависимостью.
Исходя из этого модели популяционных процессов (микроэволюционных) возможно контролировать лишь с учетом влияния филогенетической близости видов животных либо растений.
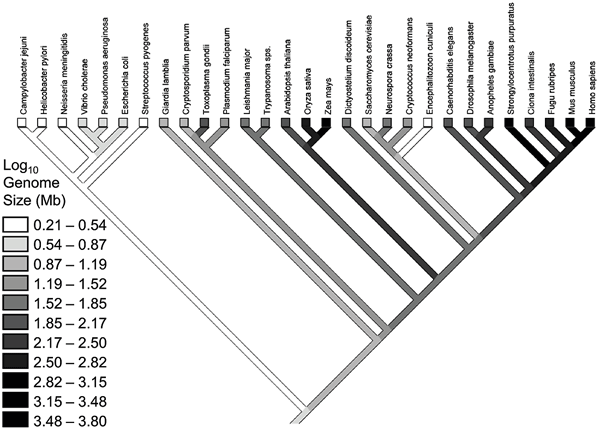
![]()
Рис.
2. филогенетическое древо и Набор видов для них с оценками размера генома, использованные американскими учеными. Изображение из обсуждаемой статьи в PLoS Genetics
На рис. 2 отлично заметно повышение размеров генома в ходе эволюции. есть ли размер популяции ведущим причиной этого повышения? Нет, не есть, заключили исследователи.
Им не удалось распознать значимую корреляцию между размером популяции и самыми различными чертями геномов: числом генов, числом и размером интронов, скоростью дупликации генов, относительным и полным числом транспозонов.
Так, разбирая два изучения, совершённые на противоположных финишах света, приходим к заключению, что мы пока не можем (к сожалению!) осознать сути геномных процессов, управляющих разрастанием либо сжатиемя геномов во времени. Мы лишь можем зафиксировать явление — рост размеров «нужной» части генома в некоторых эволюционных линиях, увязанный каким-то непонятным образом с возрастанием сложности морфофизиологической организации, — и констатировать его ускоренный темперамент. И на каких-то дорожках этого запутанного изучения мы уже можем поставить символ «тупик», дабы не тратить деньги и время на ненужные маршруты.
Источники:
1) Kenneth D. Whitney, Theodore Garland, Jr. Did Genetic Drift Drive Increases in Genome Complexity? // PLoS Genetics. 2010. 6(8): e1001080.
Doi:10.1371/journal.pgen.1001080.
2) A. В. Марков, В. А. Анисимов, А. В. Коротаев. Связь размера генома и сложности организма в эволюционном последовательности от прокариот к млекопитающим // Палеонтологический издание. №4, 2010. Стр. 3–14.
Полный текст — PDF, 207 Кб.
Елена Наймарк